


Photosynthese
Die Geschichte der Photosynthese
Wichtige Erforscher der Photosynthese und ihrer einzelnen Schritte waren:
Joseph Priestley 1733-1804
Er führte 1771 die entscheidendsten Versuche durch, die erkennen ließen, daß Pflanzen, die sogenannte " gute Luft ", welche von Tieren oder von Feuer (bei Verbrennung) verbraucht wurde herstellen. Er führte dazu zwei wichtige Versuche durch:
- Er ließ in einem geschlossenen Raum die Luft durch
eine Kerze und später durch ein Tier verbrauchen. Danach
setzte er in diese " verbrauchte " Atmosphäre
eine Pflanze, nach weiteren zehn Tagen wieder eine Kerze beziehungsweise
ein Tier.
Beobachtung:
Das Tier konnte nun darin wieder leben beziehungsweise die Kerze brennen. - Er stellte nun die Wechsel Wirkung zwischen Triumph und Pflanzen fest, indem er beide zusammen in einen Behälter gab und aufzeigen, daß Pflanzen in " verbrauchter " Luft zusammen mit einem Tier sogar besser gedeihen als in " unverbrauchter " Luft. Er schloß daraus: sowohl Tiere, die Verbrennungen verbrauchen ein Teil der Luft (" gute Luft ") und stellen dabei einen neuen das Vermögen Stoff, die " verbrauchte Luft " her. Gleichzeitig benötigen die Pflanzen dieses " verbrauchte " Luft und wandeln sie in " gute " Luft um.
Jan Ingenhousz 1730- 1799
Er entdeckte 1779 dass
- nur die grünen Teile der Pflanze zur Photosynthese dienen und
- dass dazu Licht notwendig ist.
Antoan Lavoisier 1743-1794
Er erkannte, daß der entscheidende Anteil der " verbrauchten " Luft das CO2 ist, in der " guten " Luft das O2
Jean Senebier 1742- 1809
Er erkannte 1783, das grüne Pflanzen CO2 aufnehmen.
Nicola de Saussure 1767-1845
Er wies 1804 nach, dass
- die Pflanzen das CO2 in ihrer Substanz einbauen (Substanzvergrößerung) und
- dass dabei Wasser benötigt wird.
Julius Sachs 1832-1897
Er konnte 1862 nachweisen, daß bei der Photosynthese in den Chloroplasten Stärke gebildet wird.
Robert von Mayer 1814-1878
Er erkannte bereits vorher 1845, daß bei der Photosynthese die Lichtenergie in chemische Energie umgewandelt wird, indem energiearme Stoffe (CO2, H2O) in energiereiche Stoffe (Kohlenhydrate) umgewandelt werden und in der Pflanze gespeicherten werden.
George Engelmann 1809-1884
Er führte 1882 seinen berühmten engelmannschen Bakterien- Versuch durch und erkannte damit die Hauptaktivität des sichtbaren Lichtes im blauen Wellenbereich des Spektrums 400-450 nm und im roten Wellenbereich 600-650 nm.
Blackman
Er führte die entscheidendsten Versuche zur Erkenntnis des biochemischen Vorgangs der Photosynthese 1905 durch. Er erkannte als erster, daß die Photosynthese aus zwei unabhängigen Reaktionsschritten besteht.
- Lichtreaktion (photochemische Reaktion), die Lichtabhängigkeit, aber temperaturunabhängig ist.
- eine Dunkelreaktion (normale chemische Reaktion), die lichtunabhängig, aber temperaturabhängig ist.
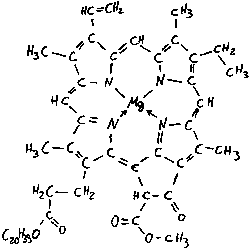
Hans Fischer 1881-1945
Er klärte die Struktur des Chlorophyllmoleküls auf (Nobelpreis 1930).
1940 wurde durch Isotopeneinsatz mittels der Tracermethode nachgewiesen, daß der bei der Photosynthese abgegebene Sauerstoff aus dem H2O stammt.
Versuch von Engelmann
Daß Licht verschiedener Wellenlänge unterschiedliche Wirkung auf die Photosynthese hat, konnte George Engelmann durch seinen Bakterienversuch beweisen. Er projizierte ein durch ein Prisma erzeugtes Licht auf einen Algenfaden. Je stärker die photosynthetische Leistung in einem Spektrum war, desto mehr Sauerstoff entstand an diesem Abschnitt und je mehr sauerstoffliebende (aerobe) Bakterien sammelten sich dort (--> Tyndalleffekt). Er fand heraus, daß im blauen und roten Strahlungsbereich ein Absorbtionsmaxima von Chlorophyll existiert. Bestrahlt man Pflanzen mit Licht aus verschiedenen Wellen und bestimmt die aus der gebildeten Sauerstoffmenge die photosynthetische Leistung, so enthält man ein Wirkungsspektrum der Photosynthese, wobei es mit dem Absorptionsspektrum der Chlorophylle übereinstimmt.
Versuch von Blackman
1905 untersuchte der Engländer Blackman die Einwirkung verschiedener Lichtstärken auf die Photosynthese bei:
- konstanter Temperatur:
Ergebnis: Bei wachsender Lichtintensität stieg die Photosyntheseleistung bis zu einem Höchstwert(Lichtsättigungspunkt) - bei Schwachlicht hat die Temperatur nur geringen Einfluß
- bei starkem Lichteinfluß steigt die Photosyntheserate dagegen mit der Temperatur an.
Bei den lichtunabhängigen chemischen Reaktionen steigt die Reaktionsgeschwindigkeit bei einer Erhöhung um 10° um das Doppelte.
Die lichtabhängigen chemischen Reaktionen sind temperaturunabhängig.
Grüne Teile der Pflanze und Blätter als Träger der Photosynthese
Wie oben dargestellt sind also die Chloroplasten die materiellen Träger der Farbstoffe, welche die Lichtreaktion der Photosynthese bewerkstelligen. Diese befinden sich in allen Zellen der grünen Teile der Pflanzen. Hauptanteil besitzen aber die Blätter der Pflanzen, welche durch ihren Bau besonders ausgebildete und für die Photosynthese besonders geeignete Organe sind. Sie sind deswegen besonders geeignet, da Sie
- folienartig aufgebaut sind, das heißt extrem dünn bei großer Oberfläche
- in ihren Zellschichten große Hohlräume zur Gas-, Wasserdampf- und Wasserspeicherung besitzen
- auf der Unterseite, das heißt der direkten Sonneneinstrahlung entzogen schlitzförmige, veränderbare Poren, Spaltöffnungen oder Stomata genannt, besitzen, welche für die Wasserabgabe und damit für den Wassersog und gleichzeitig auch für den Gasaustausch verantwortlich sind.
Zur Versteifung sind die Blätter außerdem von den Leitbündeln (s .u.) durchzogen.
Die Blätter sind wie die obige Abbildung zeigt aus nur wenigen Zellschichten aufgebaut. Die Zellen der einzelnen Zellschichten sind entsprechend ihrer Aufgaben gebaut.
- Epidermis der Oberseite: besteht aus großen unregelmäßig geformten plattenartigen Zellen welche zur Unterseite hin mit einer dicken Zellmembran ausgestattet sind und über die zusätzlich zum weiteren Verdunstungsschutz eine Kutikula gelegt ist, welche aus verschiedenen Stoffen bestehen kann, deren Grundstoff immer Zellulose ist.
- Darunter liegt eine, ein oder zwei Zellschichten dicke Palisadenschicht. Sie besteht aus zapfenförmigen, palisadenförmigen Zellen, welche sehr viele Chloroplasten besitzen. Hier erfolgt die Haupttätigkeit der Photosynthese. Die Zellen sitzen eng aneinander im Unterschied zu den Zellen des darunterliegenden Gewebes. Dieses heißt
- Schwammgewebe. Es besteht aus kleineren unregelmäßig geformten Zellen, welche senkrecht zu den palisadenförmigen liegen und damit parallel zur Blattader laufen. Sie enthalten ebenfalls Chloroplasten, aber nur in sehr geringen Mengen im Vergleich zum Palisadengewebe. Sie sind so angeordnet, das große Hohlräume entstehen, so daß der Gasaustausch gut möglich ist und sich bis zum Palisadengewebe vollziehen kann.
- Daran schließt an der Unterseite des Blattes ebenfalls eine Epidermis an, welche als einzigen Unterschied zur Oberseite Stomata enthält. Die Zellen der Stomata enthalten ebenfalls Chloroplasten.
- In das Schwammgewebe eingelagert und ebenfalls parallel zur Blattfläche verlaufend sitzen die Leitbündel. Diese bestehen auf der Oberseite aus einem wasserleitenden Teil (Xylem) und im unteren Bereich aus dem Siebteil oder Phloem.
- Der Holzteil besteht aus toten langgestreckten
Zellen, deren Zellwände am Ende aufgelöst sind, so daß
sie lange Röhren werden. Zellwände aus Zellulose sind sehr
dick und zur Versteifung mit netz-, ring- und spiralförmige Auflagerungen
aus Zellulose versehen. Der Siebteil besteht aus dünnwandigen
lebenden Zellen, deren Ende an den Enden siebartig mit Poren versehen
sind und welche die Assimilate mit dem Wasserstrom von den Blättern
in den Pflanzenkörper transportieren.
Innerhalb dieser Schicht liegen einzelne spindelförmige Zellen, welche teilungsfähig bleiben und vom Kambium des Stengels abstammen. Außerdem sind beide Teile zusammen von einem Festigungsgewebe umgeben, welche aus toten dickwandigen und stark verholzten Zellen, den Sklärenchymfaser, bestehen.
Der biochemische Vorgang der Photosynthese
Die grundlegenden Erkenntnisse für den Vorgang der Photosynthese schaffte Blackman mit seinen verschiedenen Versuchen. Aus diesen Versuchen ergibt sich:
- Es müssen zwei Redaktionen vorhanden sein, wobei die eine der RGT-Regel folgt und damit eine normale chemische Reaktion darstellt. Die andere ist dagegen eine photochemische (lichtabhängige und gleichzeitig temperaturunabhängige) Reaktion.
- Diese photochemische Reaktion ist allerdings abhängig von der Menge der Chloroplasten und Energieüberträgermoleküle, so daß sie nicht beliebig bis ins unendliche gesteigert werden kann (Lichtsättigungspunkt ).
- Auch die Temperatur kann nicht beliebig gesteigert werden, sondern hat bei 35 bis 38 Grad das Temperaturoptimum. Bei Temperaturen die über dieses Temperaturoptimum hinausgehen werden die beteiligten Enzyme unwirksam.
In den Chloroplasten liegen in den Membranen der Thylakoiden der Grana die Farbstoffe vor, während die für Photosynthese nötigen Enzyme sich in der Matrix (Stroma = Innenraum der Chloroplasten) befinden. Daher können auch die isolierten Chloroplasten photosynthetisch tätig sein. Nach heutigen Vorstellungen wird die Photosynthese in folgende Teilreaktionen eingeteilt:
- eine Lichtreaktion mit
- Bildung von ATP (Photophosphorilierung) und
- Wasserspaltung (Photolyse des Wassers)
- die Dunkelreaktion.
Photophosphorilierung
Die Chlorophyllmoleküle werden durch sichtbares Licht zum Teil in einen höheren Energiezustand versetzt, zum Teil reflektieren sie das Licht. Der Energiezustand bedeutet, daß freibewegliche Elektronen mit Photonen der gleichen Energiestufe in Resonanz treten können und damit auf ein höheres Energieniveau gehoben werden. Solche freibewegliche Elektronen liegen in Verbindung mit angegeben Gewinnung und bei speziellen Atomen vor. Beide Voraussetzungen sind bei Chlorophyllmolekülen erfüllt:
- ein Mg-Zentralatom und
- ein Porphyrinring mit konjugierter Doppelbindung.
Ein einziges Chlorophyllmolekül besitzt also zwei Elektronen im höheren Energiezustand, die leicht abgegeben werden können. Das Chlorophyll wird durch Abgabe dieser Elektronen zu Chlorophyll ++.
Die abgegebenen Elektronen werden über das eisenhaltige Protein Ferreotoxin in weitere enzymatischen Redoxsysteme an das ADP abgegeben, welches ein höherenergetisches Elektron eingebaut und dadurch befähigt wird eine weitere Phosphatgruppe zu binden und somit zum ATP zu werden. Gleichzeitig gibt das ADP ein Elektron niederen Energieniveaus an das Chlorophyll + zurück, so daß dieses wieder neutral wird und zu erneuter Reaktion fähig ist (zyklische Phosphorilierung).
Die Photolyse des Wassers
Gleichzeitig kann eine weitere andere Reaktion ablaufen. Das Elektron des Chlorophyllmoleküls kehrt nicht immer im Zug der zyklischen Photophosphorilierung wieder zum Chlorophyll zurück, sondern wird über Ferredoxin und über weitere Redoxsysteme völlig an das NADP abgegeben, an dem es bleibt. Nun fehlt aber diesem Chlorophyllmolekül das Elektron. Es holt sich dieses von einem zweiten Chlorophyllmolekül, welches sein ebenfalls angeregtes Elektron über das Plastochinon und anschließende Enzymkomplexe an das Chlorophyllmolekül 1 weiterleitet. Nun fehlt aber diesem Chlorophyllmolekül 2 das Elektron und dieses Elektron wird nun dem H2O entzogen, wobei H +-Ionen von Sauerstoff in Gasform frei werden. Die Energie zu dieser H2O-Spaltung wird einerseits ans vorhandene ATP-Molekül und andererseits aus den anderen exerkonisch ablaufenden Reaktionen der elektronen-transportierenden Redoxsysteme gewonnen.
Die freiwerdenden H +-Ionen werden an das NADP gebunden, daß bei der ersten Lichtreaktion ein angeregtes Elektron vom Chlorophyll 1 erhalten hat. Dieses daraus entstehende NADPH wird als Reduktionsmittel in der Dunkelphase benötigt. Da also das ursprüngliche Elektron vom Chlorophyll 1 beim N ADPH bleibt und das Chlorophyll 1 ein neues Elektron vom Chlorophyll 2 erhält nennt man diesen Vorgang nichtzyklische Phosphorilierung.
Die Dunkelreaktion
Die in der Lichtreaktion gewonnene Energie die in den beiden Molekülen ATP und NADPH gespeichert ist wird nun in der Dunkelreaktion zum Aufbau energiereicher Moleküle genutzt. Dabei geschieht folgendes:
Das CO2 wird nicht wie die einfache Summengleichung der Photosynthese nämlich ( 6H2O + 6CO2 ® C6H12O62 G= + 2875 kJ) vermuten ließe mit Wasser zu Zucker zusammengesetzt. Das CO2 wird vielmehr an einem C5-Körper gebunden, so daß ein C6- Körper entsteht.
Aus einem Ribolosediphosphat wird so ein C6-Körper, welcher allerdings nicht stabil ist, sondern sofort in 2 C3-Körper (Glycerinsäurephosphate) zerfällt. Diese Körper werden mit Hilfe des Wasserstoffs, welcher vom NADPH stammt zu Glycerinaldehydphosphat reduziert (= Triosephosphat). Dazu wird außerdem ATP benötigt, welche die Energie für diese Umsetzung liefert. Diese beiden Glycerinaldehydphosphate reagieren dann miteinander zu einem C6-Körper, dem Fructosediphosphat. Davon wird zunächst ein
P-Rest abgespalten und daraus ein Glucosemonophosphat herstellt. Daneben kann es auch Fructosemonophosphat bleiben (Herstellung von Saccharose). Dies geschieht allerdings nur mit jedem sechsten C-Atom. Die übrigen fünf Fructosemonophosphate wandeln sich über diverse Zwischenschritte wieder in einen C5-Körper, nämlich das Ribolosediphosphat, um, daß dann erneut für eine CO2-Anlagerung zur Verfügung steht.
Der gesamte Vorgang wird auch Calvin-Benson-Zyklus genannt.
Die so gewonnenen Zucker Glucose beziehungsweise Fructose werden entweder gespeichert (Stärke) oder als löslicher Zucker (Saccharose = Zweierzucker aus Fructose und Glucose) in das Plasma der Zelle beziehungsweise die Zellsafträume der Pflanze gebracht, wo er als Nährstoffe/Baustoff oder wiederum Speicherstoff zur Verfügung steht.
Materielle Träger der Photosynthese sind die Chloroplasten (vgl. Buch S. 126). In den Chloroplasten liegen also die Blattfarbstoffe in den Thylakoiden in der Membran, besonders in dem Grana- Thylakoid vor. Die Enzyme für die Abläufe der Dunkelreaktion liegen dagegen in der Matrix. Da zusätzlich in den Chloroplasten DNA und Ribosomen vorhanden sind, können auch isolierte Chloroplasten photosynthetisch tätig sein. Insgesamt gibt es folgende Blattfarbstoffe:
- Chlorophyll a
- Chlorophyll b
- Xanthophylle
- Carotine
Letztere zusammen werden auch Carotinoide genannt.
Allen diesen Farbstoffen ist gemeinsam, dass sie freie delokalisierte p- Elektronen besitzen. Das freie p- Elektronensystem beruht auf einem Porphyrinringsystem, dass der Carotinoide auf einen langkettigen, linearen Elektronen- System (40 C- Atome) und das der Xantrophylle aus einem Ringsystem bestehend aus 2 Ringen mit n- Atomen und Seitenketten mit Doppelbindungen.
